 腾讯登录
腾讯登录科研 | 剪接体全体结构破译指日可待 ——施一公团队再发Science解析完全组装剪接体激活前结构
| 导读 | 完全组装的酿酒酵母剪接体激活前结构 |
导读
大家都在高中生物课上学过遗传信息传递的中心法则——DNA被转录生成信使RNA,信使RNA被翻译成蛋白质(其中DNA和RNA都可以自我复制)。如果你进一步学过分子生物学或生物化学,知道真核生物DNA基因的不连续性,它们包括编码蛋白的外显子和不编码蛋白的内含子。因此,遗传信息从DNA到RNA(称pre-mRNA,前体信使RNA)后,需要经历内含子剪除和外显子拼接,这一过程被称为RNA剪接。而剪接体(spliceosome)是RNA剪接的主要执行者,它们由多种蛋白质和RNA组成(ribonucleoprotein,RNP),识别pre-mRNA上5‘剪接位点,按照高度精确的顺序发生不同组分的复合物组装、激活、催化、转脂等过程,最终解离成小的结构单元,进入下一个循环反应。目前已知的剪接体复合物依次为E、A、pre-B、B、Bact、B*、C、C*、P、ILS(红色表示结构已知)等。这篇论文报道的是激活前的两个完全组装结构,称为预催化剪接体前体pre-B复合物和预催化剪接体B复合物,分辨率分别为3.3-4.6埃和3.9埃。该结构是目前组分最多、分子量最大的剪接体结构,清晰显示了剪接体组装前期pre-mRNA的5‘剪接位点识别状态和动态变化,为解释剪接体如何逐步组装和激活提供了至关重要的结构基础。

论文ID
原名:Structures of the fully assembled Saccharomyces cerevisiae spliceosome before activation
译名:完全组装的酿酒酵母剪接体激活前结构
期刊:Science
IF:37.2
发表时间:2018年5月
通信作者:施一公
通信作者单位:清华大学

实验内容
1、pre-B和B复合物冷冻电镜结构
pre-B复合物结构(图1)是目前已知的组分最多、分子量最大的剪接体,由U1 snRNP(包含16个蛋白)、U2 snRNP(包含18个蛋白)、tri-snRNP(U4,U5,U6,31个蛋白)和RES(包含3个蛋白)四个复合物组装形成,共有68个蛋白和6条RNA(5条snRNA,一条pre-mRNA)。B复合物共55个蛋白(图1B),4条snRNA和1条pre-mRNA。这两个结构首次清楚地展示了剪接体组装前期U1 snRNP识别的pre-mRNA5‘剪接位点,以及各组分蛋白、RNA间的相互作用。
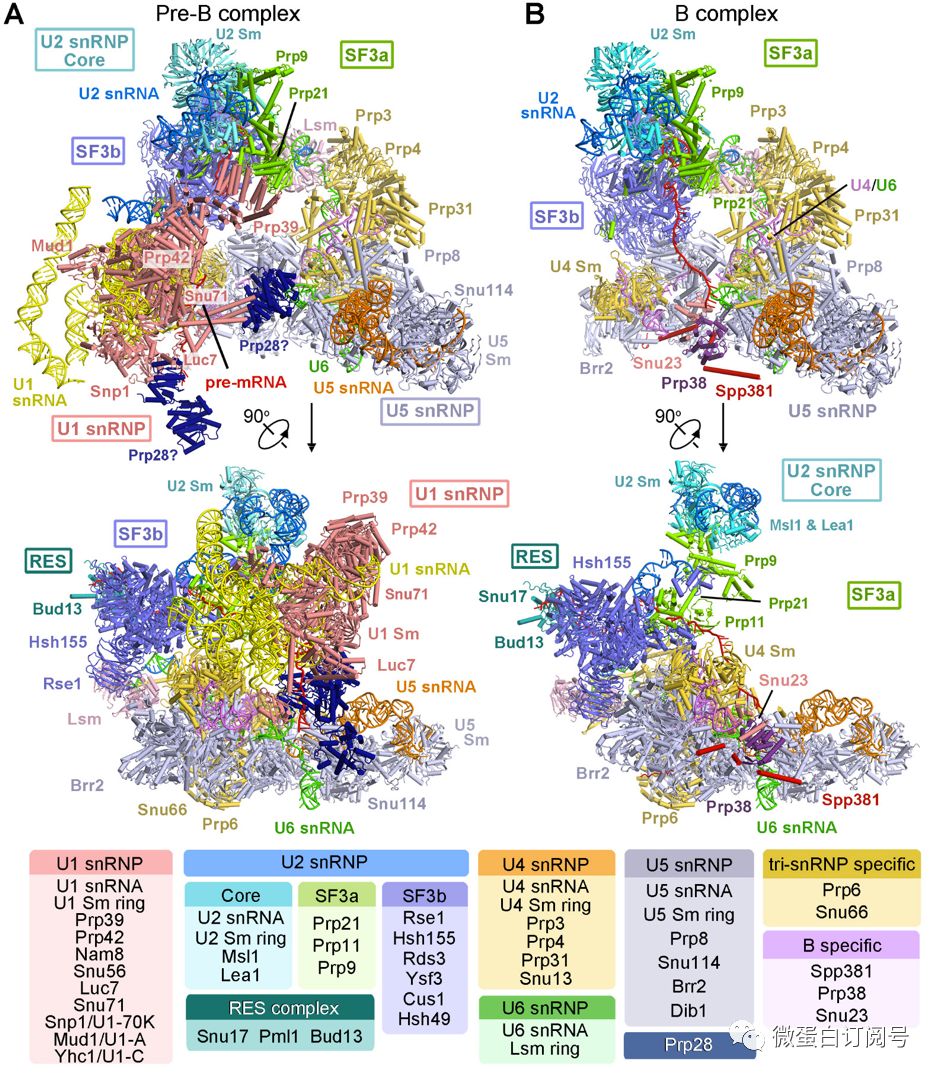
图1 完全组装的预催化剪接体前体Pre-B和预催化剪接体B复合物结构。
2、pre-B向B转变的结构改变
pre-B与B复合物结构区别主要包括:(1)B复合物没有U1 snRNP组分(图2A中粉色),这使得1a)pre-mRNA5‘剪接位点由起初的被U1 snRNP识别,转为被U6 snRNP识别;1b)pre-B中原有的U1 snRNP和U2 snRNP相互作用消失,该相互作用由U2 snRNA和U1 Prp42蛋白表面带正电的残基维系(图2B);(2)B复合物中U2 snRNP转向tri-snRNP,两组分相互作用更强,这使得2a)tri-snRNP组分中的Brr2蛋白旋转了~30度,空间位置转移了40-50埃(图2E);2b)U2 snRNP组分中的Hsh155蛋白移向tri-snRNP组分中LSm蛋白(图2C和2F中黄色与淡粉色),U2 snRNP中的Cus1与Rse1蛋白和tri-snRNP中的Brr2蛋白由原来的无相互作用(图2D)到有直接接触(图2G);(3)B复合物中新招募了三个蛋白(Prp38、snu23和Spp381),且三者形成相互作用(图2H)。其中,Prp38和Snu23的表面带正电势能可能对pre-mRNA的5’末端摆向有一定贡献(图2I)。
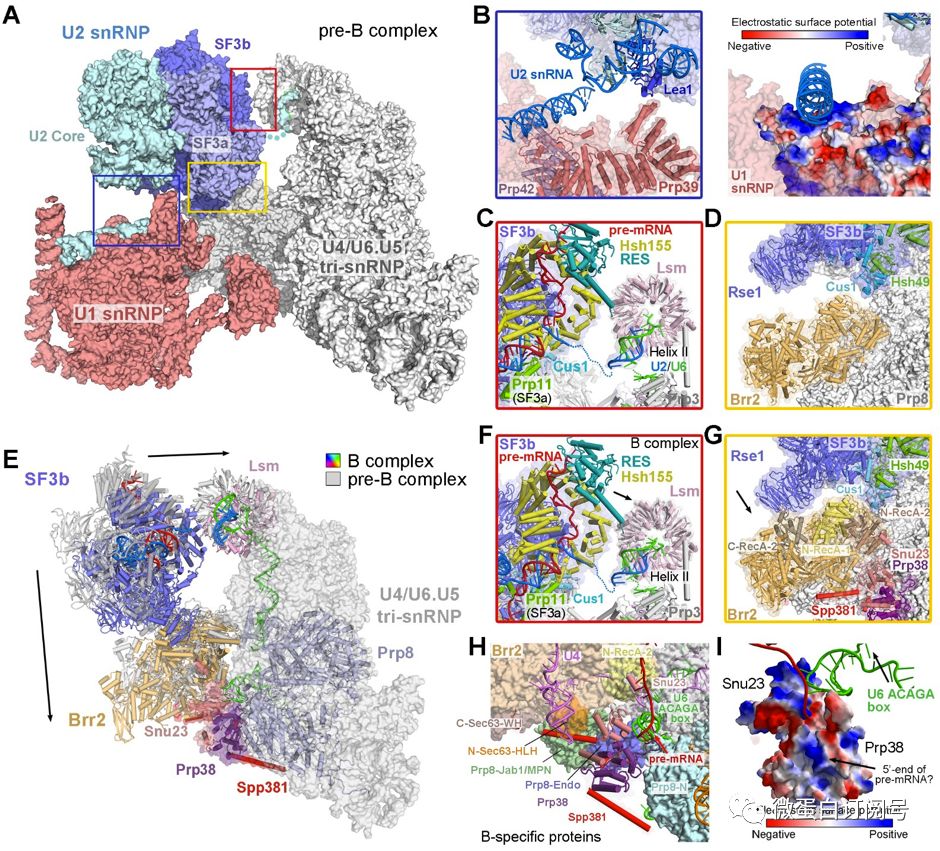
图2 pre-B和B复合物结构的改变及对具体组分构象的影响
3、不完全组装的预剪接体A复合物结构模型
由于pre-B复合物是由A复合物(包含U1和U2-snRNP)和tri-snRNP组装而成,本文解析的pre-B中U1和U2-snRNP只是短暂地与tri-snRNP形成接触界面,因此A复合物中的U1和U2-snRNP构象可能与pre-B中的U1和U2-snRNP相似。根据这个推测,本文构建了更早期的不完全组装预剪接体A复合物模型(图3A)。
之前的研究工作表明,剪接体组装第一步是A复合物结合tri-snRNP形成pre-B复合物;第二步,在腺苷三磷酸酶/解旋酶Prp28的作用下,U1-snRNP解离,释放的5‘剪接位点在B复合物特异蛋白的帮助下重置于U6和U5 snRNA附近;第三步,在腺苷三磷酸酶/解旋酶Brr2的作用下,B复合物经历构象变化,形成Bact复合物(图3B)。
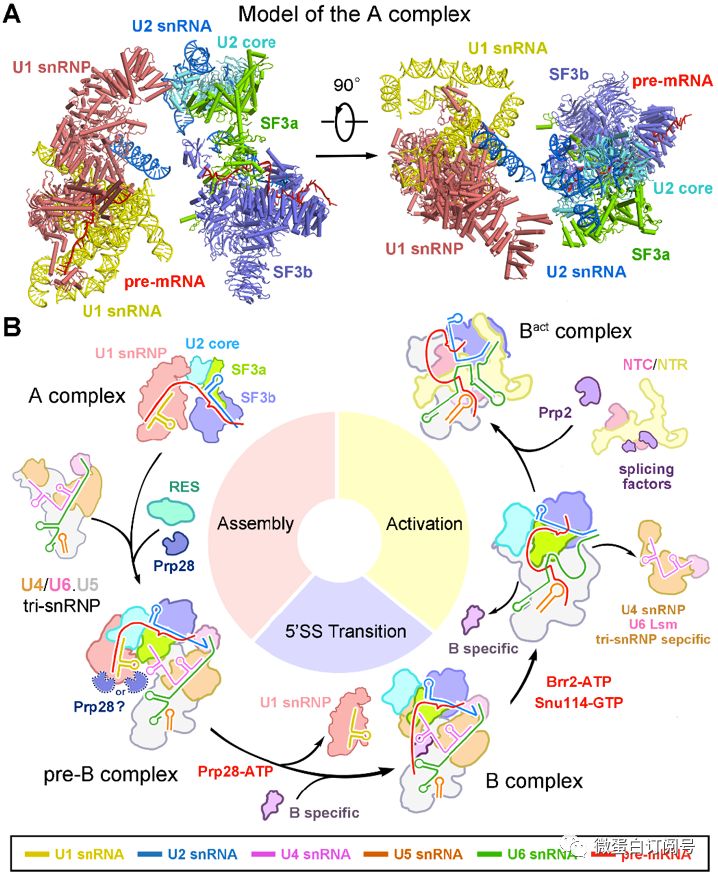
图3酿酒酵母剪接体组装和激活机制。(A)推测的不完全组装复合物A结构模型。(B)酿酒酵母剪接体组装和激活机制流程图。
 实验结论
实验结论
本次报道的结构,和之前不同状态的剪接体结构贯连在一起,清楚地展示了pre-mRNA在剪接体组装和激活状态下的变动。具体地,从pre-B到B,pre-mRNA5‘剪接位点重置了~90埃,从B到Bact,重置了40埃。
RNA剪接的每一过程都由特定RNA和蛋白质组装而成的复合物参与,施一公团队自2015年8月报道首个裂殖酵母剪接体冷冻电镜结构(包含U2,U6,U5,NTC,NTR,位于图4中央)以来,共解析了剪接过程的9个高分辨率复合物结构(图4),包括预组装复合物U4/U6.U5 Tri-snRNP、激活状态的Bact、第一步催化反应后复合物C、第二步催化激活状态下的C*、转脂反应后的P复合物、内含子套索剪接体ILS复合物,以及本篇论文解析的完全组装激活前pre-B和B复合物。至此,完全组装的剪接体结构中,只剩下B*复合物结构未知。
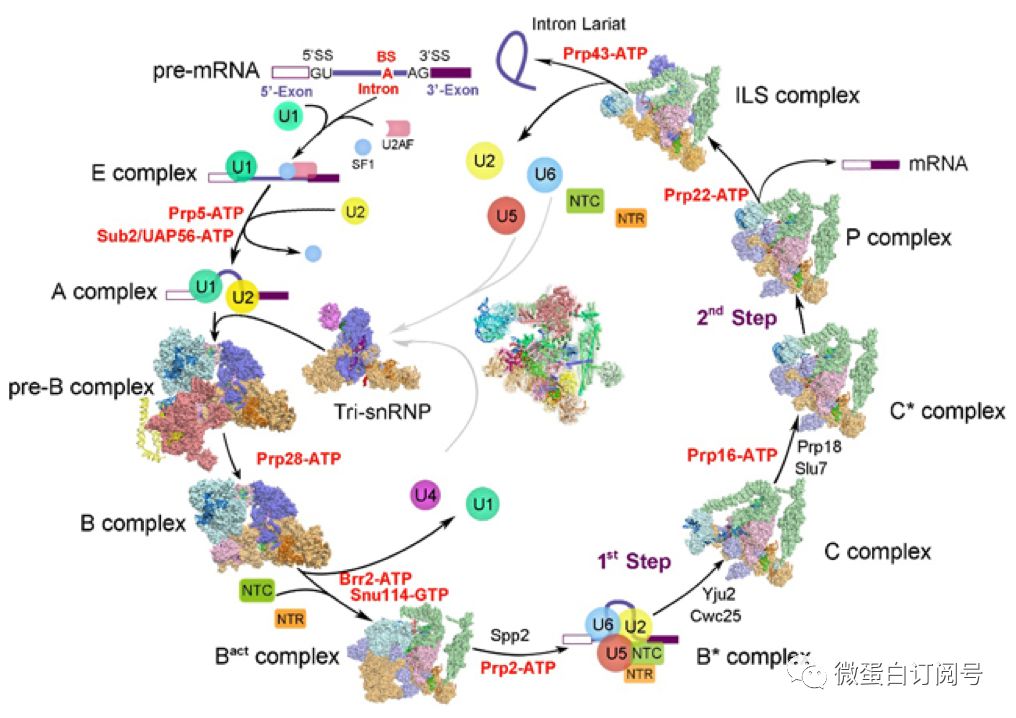
图4 施一公团队解析的剪接体结构汇总
点 评
相信在不久的将来,图4中的所有复合物结构都会问世,这些结构的解析,将清楚地呈现出剪接体组装前后、激活前后、催化反应前后、转脂反应前后构象的变化,为揭示剪接体分子机制提供最直接全面的结构基础,对剪接体研究领域做出极为重要的贡献。
(转化医学网360zhyx.com)
还没有人评论,赶快抢个沙发