 腾讯登录
腾讯登录CTLA4
| 导读 | 概述
1型糖尿病(DM)是免疫介导的胰岛细胞损伤而引起的自身免疫性疾病。许多研究表明,T细胞介导了免疫损伤过程。虽然所涉及的免疫病理机制尚不清楚,但是近年来通过对CD28细胞毒性T淋巴细胞相关抗原(CTLA)-4-B7分子的深入研究,证明其在1型糖尿病的发生、发展中起着重要作用。
T细胞的活化需要双信号,T细胞通过其抗原受体(TCR)... |
概述
1型糖尿病(DM)是免疫介导的胰岛细胞损伤而引起的自身免疫性疾病。许多研究表明,T细胞介导了免疫损伤过程。虽然所涉及的免疫病理机制尚不清楚,但是近年来通过对CD28细胞毒性T淋巴细胞相关抗原(CTLA)-4-B7分子的深入研究,证明其在1型糖尿病的发生、发展中起着重要作用。
T细胞的活化需要双信号,T细胞通过其抗原受体(TCR)识别抗原递呈细胞(APC)表达的主要组织相容复合物(MHC)后,即获得第一信号,诱导T细胞表达CD40L,与APC持续表达的CD40相互作用,使APC上调性表达B7分子。B7分子与T细胞的CD28相互作用,使T细胞获得辅助刺激信号(第二信号),导致T细胞活化,分化成有功能的效应T细胞,从而诱导其合成、分泌细胞因子,同时诱导其他共刺激分子如CD40L分子的表达。CD40与CD40L的相互结合又诱导B7的上调性表达,进一步上调T细胞的活性[1, 2]。
研究表明,T细胞必须在第一信号及辅助刺激信号共同作用下才能完全活化。前者是T细胞活化的基础,并决定了T细胞反应的特异性;后者是T细胞活化的必要条件,两者缺一不可。缺乏或阻断辅助刺激信号,将会引起T细胞的无反应性或细胞凋亡及克隆丢失[3]。后者为T细胞膜上表面分子,前者为表达于特异性抗原递呈细胞的膜上的相应配体,二者相互作用发挥辅助刺激作用。
基因结构
CTLA4(Cytotoxic Lymphocyte Antigen 4,CTLA4)是细胞毒性T淋巴细胞中发现的第四种特异性抗原基因。该基因位于人类第2号染色体的2q33位置,基因全长6,174bp,含有3个外显子和2个内含子。
基因分子生物学功能
CTLA4蛋白的胞外段的主要功能是与其相关配体B7分子结合,现在已发现三种以上的B7分子,分别命名为B7-1(CD80)、B7-2(CD86)、B7-3等。CTLA4胞外区与B7分子的结合力比CD28大20至100倍[4]。CD28是一分子量为44KD的同源二聚体糖蛋白,由二硫键连接,主要表达于T细胞和浆细胞表面,该蛋白和CTLA4蛋白的结构具有同源性,都作用于同样的配体,但是产生的作用却不同。CD28活化介导的共刺激通路正调节免疫反应,CTLA4蛋白则对免疫反应是负性调节[5]。CTLA4蛋白的跨膜区是将胞外区被B7分子结合的信号传向细胞内,CTLA4胞内区是将外来信号转变成细胞的负调信号,从而降低T细胞功能。
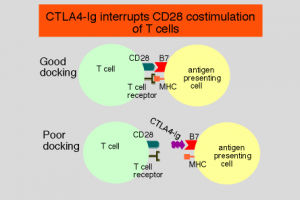
如图1所示,CTLA4基因编码的CTLA4蛋白一方面可竞争性地与特异性抗原递呈细胞膜上的B7分子结合,从而阻断T淋巴细胞活化中的B7-CD28辅助信号途径,进而阻断T细胞的活化;另一方面,当CTLA4分子与B7分子结合后,致CTLA4胞内段磷酸化,产生负调信号,从而阻断T淋巴细胞活化及功能,因而认为CTLA4是一种T细胞活化负调节因子。
众多研究表明,CTLA4阻断B7-CD28辅助信号途径在下调T细胞功能中起着重要作用。目前的研究证实,CTLA4基因多态能改变CTLA4与B7分子结合的活性,从而增加了相关免疫疾病发生的危险度。参与的通路
CTLA4蛋白在体内主要参与细胞免疫应答。如图2所示,该过程从T细胞的TCR识别APCs的MHC-抗原肽复合物,并激活T细胞开始的。TCR复合体是由参与抗原识别的T细胞受体α、β链异二聚体与γδεζ链组成的CD3复合体偶联形成的。TCR的α、β链因胞内区较短,无信号传导功能,信号传导主要由胞内区较长的CD3分子完成。
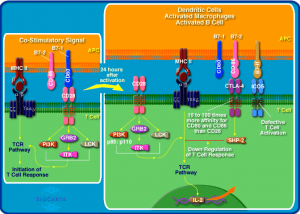
在识别抗原时,TCR结合抗原后招募CD3分子,首先发生CD3分子(特别是CD3的ζ链)的酪氨酸磷酸化,CD3分子具有胞内信号基序ITAM(im-munoreceptor tyrosine-based activation motif)。ITAM是YXXL/I序列间隔有11个氨基酸的二次重复结构,伴随着T细胞的活化,该基序中的酪氨酸被Src激酶家族Lck、Fyn磷酸化。Lck作为辅助受体,与CD4/CD8结合,而Fyn直接与TCR复合体结合。酪氨酸磷酸化的ITAM可招募Syc酪氨酸激酶家族的ZAP-70。ZAP-70通过并列排列的2个SH-2区与2个磷酸化YXXL/I结合,发生自身磷酸化,也可被Lck磷酸化后而激活,活化的ZAP-70可诱发下游分子发生磷酸化并引发一系列信号传导。
在免疫应答的过程中,CTLA4至少通过两条途径发挥着下调T细胞功能的作用:一方面是通过CTLA4胞外区和CD28竞争性结合,而抑制抗原细胞上的B7分子与T细胞膜上的CD28分子结合,从而阻断T细胞活化所必须的B7-CD28辅助信号途径,致使T细胞功能下调;另一方面,当CTLA4与B7分子结合后,直接产生负调信号,同样下调T细胞功能。
研究表明,CTLA4蛋白在T细胞活化初期并不参与负调节,随活化的延长才开始被诱导表达,主要是防止超敏免疫反应的发生,并诱发抑制性信号最终导致免疫反应终止,这可能就是其负调节的机理。CTLA4介导的抑制性反应在抗原特异性T细胞活化时,加入抗CTLA4抗体可解除原来的抑制反应,并增强T细胞活化。
基因对疾病发生的影响
CTLA4基因变异导致CTLA4蛋白与B7蛋白分子的结合力下降,使得CTLA4蛋白对免疫信号的下调作用降低,破坏了正常的免疫信号调节,产生1型糖尿病等免疫性疾病。
1型糖尿病(DM)是免疫介导的胰岛细胞损伤而引起的自身免疫性疾病。许多研究表明,T细胞介导了免疫损伤过程。虽然所涉及的免疫病理机制尚不清楚,但是近年来通过对CD28细胞毒性T淋巴细胞相关抗原(CTLA)-4-B7分子的深入研究,证明其在1型糖尿病的发生、发展中起着重要作用。
T细胞的活化需要双信号,T细胞通过其抗原受体(TCR)识别抗原递呈细胞(APC)表达的主要组织相容复合物(MHC)后,即获得第一信号,诱导T细胞表达CD40L,与APC持续表达的CD40相互作用,使APC上调性表达B7分子。B7分子与T细胞的CD28相互作用,使T细胞获得辅助刺激信号(第二信号),导致T细胞活化,分化成有功能的效应T细胞,从而诱导其合成、分泌细胞因子,同时诱导其他共刺激分子如CD40L分子的表达。CD40与CD40L的相互结合又诱导B7的上调性表达,进一步上调T细胞的活性[1, 2]。
研究表明,T细胞必须在第一信号及辅助刺激信号共同作用下才能完全活化。前者是T细胞活化的基础,并决定了T细胞反应的特异性;后者是T细胞活化的必要条件,两者缺一不可。缺乏或阻断辅助刺激信号,将会引起T细胞的无反应性或细胞凋亡及克隆丢失[3]。后者为T细胞膜上表面分子,前者为表达于特异性抗原递呈细胞的膜上的相应配体,二者相互作用发挥辅助刺激作用。
基因结构
CTLA4(Cytotoxic Lymphocyte Antigen 4,CTLA4)是细胞毒性T淋巴细胞中发现的第四种特异性抗原基因。该基因位于人类第2号染色体的2q33位置,基因全长6,174bp,含有3个外显子和2个内含子。
基因分子生物学功能
CTLA4蛋白的胞外段的主要功能是与其相关配体B7分子结合,现在已发现三种以上的B7分子,分别命名为B7-1(CD80)、B7-2(CD86)、B7-3等。CTLA4胞外区与B7分子的结合力比CD28大20至100倍[4]。CD28是一分子量为44KD的同源二聚体糖蛋白,由二硫键连接,主要表达于T细胞和浆细胞表面,该蛋白和CTLA4蛋白的结构具有同源性,都作用于同样的配体,但是产生的作用却不同。CD28活化介导的共刺激通路正调节免疫反应,CTLA4蛋白则对免疫反应是负性调节[5]。CTLA4蛋白的跨膜区是将胞外区被B7分子结合的信号传向细胞内,CTLA4胞内区是将外来信号转变成细胞的负调信号,从而降低T细胞功能。
图1 CTLA4蛋白竞争性阻断CD28与B7蛋白的结合
如图1所示,CTLA4基因编码的CTLA4蛋白一方面可竞争性地与特异性抗原递呈细胞膜上的B7分子结合,从而阻断T淋巴细胞活化中的B7-CD28辅助信号途径,进而阻断T细胞的活化;另一方面,当CTLA4分子与B7分子结合后,致CTLA4胞内段磷酸化,产生负调信号,从而阻断T淋巴细胞活化及功能,因而认为CTLA4是一种T细胞活化负调节因子。
众多研究表明,CTLA4阻断B7-CD28辅助信号途径在下调T细胞功能中起着重要作用。目前的研究证实,CTLA4基因多态能改变CTLA4与B7分子结合的活性,从而增加了相关免疫疾病发生的危险度。参与的通路
CTLA4蛋白在体内主要参与细胞免疫应答。如图2所示,该过程从T细胞的TCR识别APCs的MHC-抗原肽复合物,并激活T细胞开始的。TCR复合体是由参与抗原识别的T细胞受体α、β链异二聚体与γδεζ链组成的CD3复合体偶联形成的。TCR的α、β链因胞内区较短,无信号传导功能,信号传导主要由胞内区较长的CD3分子完成。
图2 CTLA4基因参与的免疫调节通路
在识别抗原时,TCR结合抗原后招募CD3分子,首先发生CD3分子(特别是CD3的ζ链)的酪氨酸磷酸化,CD3分子具有胞内信号基序ITAM(im-munoreceptor tyrosine-based activation motif)。ITAM是YXXL/I序列间隔有11个氨基酸的二次重复结构,伴随着T细胞的活化,该基序中的酪氨酸被Src激酶家族Lck、Fyn磷酸化。Lck作为辅助受体,与CD4/CD8结合,而Fyn直接与TCR复合体结合。酪氨酸磷酸化的ITAM可招募Syc酪氨酸激酶家族的ZAP-70。ZAP-70通过并列排列的2个SH-2区与2个磷酸化YXXL/I结合,发生自身磷酸化,也可被Lck磷酸化后而激活,活化的ZAP-70可诱发下游分子发生磷酸化并引发一系列信号传导。
在免疫应答的过程中,CTLA4至少通过两条途径发挥着下调T细胞功能的作用:一方面是通过CTLA4胞外区和CD28竞争性结合,而抑制抗原细胞上的B7分子与T细胞膜上的CD28分子结合,从而阻断T细胞活化所必须的B7-CD28辅助信号途径,致使T细胞功能下调;另一方面,当CTLA4与B7分子结合后,直接产生负调信号,同样下调T细胞功能。
研究表明,CTLA4蛋白在T细胞活化初期并不参与负调节,随活化的延长才开始被诱导表达,主要是防止超敏免疫反应的发生,并诱发抑制性信号最终导致免疫反应终止,这可能就是其负调节的机理。CTLA4介导的抑制性反应在抗原特异性T细胞活化时,加入抗CTLA4抗体可解除原来的抑制反应,并增强T细胞活化。
基因对疾病发生的影响
CTLA4基因变异导致CTLA4蛋白与B7蛋白分子的结合力下降,使得CTLA4蛋白对免疫信号的下调作用降低,破坏了正常的免疫信号调节,产生1型糖尿病等免疫性疾病。

评论:
共有 1 条评论
-
游客2018-12-09 18:02:30图2有没有高清图?
加载更多
 正在加载
正在加载没有更早的了