 腾讯登录
腾讯登录IL1RN
| 导读 | 概述
骨重建的结果取决于骨吸收和骨形成的相互关系。破骨细胞(OC)是骨吸收的主要细胞;而成骨细胞(OB)是骨形成的主要细胞。OC和OB的数目和活性比例决定了骨重建的结果:破骨作用大于成骨作用,将造成骨质丢失、骨质疏松(OP)、甚至病理性骨折。OC参与骨吸收的过程包括OC分化、活化、降解移出骨基质和矿物质。许多研究证明IL-1在上述过程发挥重要作用。因此IL-1被称为“... |
概述
骨重建的结果取决于骨吸收和骨形成的相互关系。破骨细胞(OC)是骨吸收的主要细胞;而成骨细胞(OB)是骨形成的主要细胞。OC和OB的数目和活性比例决定了骨重建的结果:破骨作用大于成骨作用,将造成骨质丢失、骨质疏松(OP)、甚至病理性骨折。OC参与骨吸收的过程包括OC分化、活化、降解移出骨基质和矿物质。许多研究证明IL-1在上述过程发挥重要作用。因此IL-1被称为“破骨细胞活化因子”。
IL-1是能影响多种细胞的细胞因子家族,由IL-1α、IL-1β及IL-1Ra组成。IL-1Ra能竞争性地结合IL-1R,阻断IL-1α、IL-1β与之结合,但不引起目前可测的激动性效应,是天然存在的IL-1拮抗剂。
大量关联研究表明,IL-1与骨质疏松症的形成有关,IL-1激活破骨细胞使得骨质流失,而白介素1受体拮抗物(IL-1Ra)则抑制此过程。对IL1RN基因多态性的研究,也显示第2号内含子的VNTR与IL-1Ra的表达水平有关,因此也与骨质疏松症相关联。
基因结构
IL1RN全称为interleukin 1 receptor antagonist(白细胞介素1受体拮抗物),位于染色体2q14.2,全长16,123 bp。mRNA长度根据其异构体而定,分别为1,802、1,865、1,769、1,973nt。编码的蛋白也有四种长度,分别由159、180、177、143个氨基酸残基组成。
其中,编码分泌型蛋白(sIL-1Ra)和细胞型蛋白(icIL-1Ra)的基因都位于2号染色体2q13-14.1的基因复合体中,由4个共用外显子、3个共用内含子和各自特异的第一个外显子组成。sIL-1Ra基因的特异外显子处于第1个共用外显子之前,上面有一个剪接位点。该外显子被剪切后,连接上3种亚型的icIL-1Ra各自特异的外显子,便构成了icIL-1Ra的序列。进一步翻译表达就产生了带信号肽的sIL-1Ra和不带信号肽的icIL-1Ra。
基因分子生物学功能
IL-1具有广泛的生物学作用,例如:①IL-1作为内源性致热原(endogenous pyrogen)而引起炎症反应;②促进T细胞分泌IL-2和表达IL-2R,直接或间接地通过诱导Th2细胞产生IL-4、IL-5、IL-6来促进B细胞增殖和分化,参与特异性免疫应答;③促进单核/巨噬细胞及血管内皮细胞合成细胞因子、粘附分子,激活白细胞粘附作用和合成炎性蛋白;④促进肝细胞合成急性期蛋白(acute phase protein),从而参与炎症反应;⑤促进伤口愈合;⑥刺激造血;⑦直接或间接发挥杀瘤效应;⑧IL-1Ra可与IL-1竞争性结合靶细胞表面IL-1R,从而发挥抑制作用。
IL-1可看作是能影响多种细胞的细胞因子家族,由IL-1α、IL-1β及IL-1Ra组成。IL-1α、IL-1β与白介素-1受体(IL-1R)结合而发挥广泛的细胞活性激动剂的作用[1]。IL-1Ra(编码基因为IL1RN)是80年代从尿中分离出,可拮抗IL-1作用的细胞因子。其氨基酸序列与IL-1β有26%同源性,与IL-1α有19%同源性,能竞争性地结合IL-1R,但不引起目前可测的激动性效应,是天然存在的IL-1拮抗剂。目前已发现的IL-1Ra有2类4型:分泌型(sIL-1Ra)、细胞型(icIL-1Ra)Ⅰ、细胞型Ⅱ、细胞型Ⅲ[2]。sIL-1Ra主要由单核细胞、巨噬细胞、中性粒细胞等分泌,其N端含有一个25个氨基酸残基的信号肽,主要作用是直接调节免疫反应,包括自身免疫反应。icIL-1Ra存在于角质细胞、肠上皮细胞以及视网膜上皮细胞等,其N端只带有7个氨基酸残基而非信号肽,因此无法转运至胞外。icIL-1Ra在胞内缓冲IL-1的作用,并削弱IL-1诱导产生的基因表达作用[3],该蛋白还可在细胞坏死时释放到胞外发挥类似sIL-1Ra的效能[4]。
IL-1R分为Ⅰ型和Ⅱ型,均属免疫球蛋白超家族。Ⅰ型受体广泛分布于T细胞、成纤维细胞、平滑肌细胞和内皮细胞表面,是IL-1的信号传导受体。Ⅱ型受体主要分布于B细胞、中性粒细胞和单核/巨噬细胞表面。两者信号转导方式不同,具体见图1。
一般,在细胞表面有大量的IL-1R,只需要极小量(<10个)的IL-1与IL-1R结合,即可产生显著生物学效应。IL-1Ra与IL-1R结合而不产生效应是因为它缺乏与IL-1受体辅助蛋白(IL-1R2Acp)结合的区域,因此不具有进一步激发信息传导的功能。再者IL-1Ra半衰期很短(6min),且容易与血循环中可溶性IL-1R结合从而降低其他组织器官中IL-1Ra的水平,加上病变部位IL-1浓度可能很高,因此要完全拮抗IL-1的作用需要10甚至1000倍于IL-1的IL-1Ra数量才能有效。
参与的通路
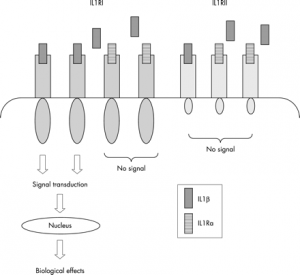
图1 IL-1R的两种受体蛋白介导的信号转导示意图
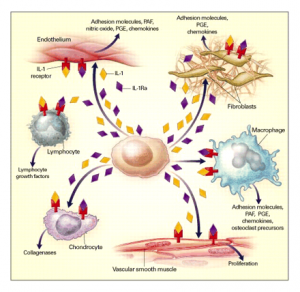
图2 IL-1和IL-1Ra的作用途径
基因对疾病发生的影响
白细胞介素-1(IL-1)是一类作用很广泛的细胞因子,参与机体的炎症、免疫防御反应、骨髓造血功能调节及几乎包括所有器官系统的自体平衡机制。其中IL-1α和IL-1β是受体激动剂,与机体免疫功能激活密切相关。研究表明,IL-1常参与各种疾病的发生发展,如败血症、自身免疫性疾病等的病理过程。过多的IL-1会加剧病情的恶化,而恶化的病理过程产生更多IL-1,由此形成恶性循环。另外的研究发现,该家族的另一成员——白介素-1受体拮抗物(IL-1Ra),可缓解此过程,打破恶性循环,因而受到极大的重视。
IL-1在骨代谢中具有双向调节作用,低浓度的IL-1可以促进骨质形成并可作为骨合成代谢的一个指标,然而高浓度的IL-1则可促进骨质丢失而成为与骨密度(BMD)成反比的生化指标。IL-1Ra是特异性与IL-1发生拮抗的细胞因子,必然同IL-1在骨质代谢中发生对抗,从而具有抗骨质疏松症的作用。
功能及人群关联性研究显示,IL1RN基因多态性,特别是研究较多的第2号内含子上VNTR,会影响IL-1Ra的表达水平,因而可能在IL-1介导的骨质流失中扮演重要角色。
骨重建的结果取决于骨吸收和骨形成的相互关系。破骨细胞(OC)是骨吸收的主要细胞;而成骨细胞(OB)是骨形成的主要细胞。OC和OB的数目和活性比例决定了骨重建的结果:破骨作用大于成骨作用,将造成骨质丢失、骨质疏松(OP)、甚至病理性骨折。OC参与骨吸收的过程包括OC分化、活化、降解移出骨基质和矿物质。许多研究证明IL-1在上述过程发挥重要作用。因此IL-1被称为“破骨细胞活化因子”。
IL-1是能影响多种细胞的细胞因子家族,由IL-1α、IL-1β及IL-1Ra组成。IL-1Ra能竞争性地结合IL-1R,阻断IL-1α、IL-1β与之结合,但不引起目前可测的激动性效应,是天然存在的IL-1拮抗剂。
大量关联研究表明,IL-1与骨质疏松症的形成有关,IL-1激活破骨细胞使得骨质流失,而白介素1受体拮抗物(IL-1Ra)则抑制此过程。对IL1RN基因多态性的研究,也显示第2号内含子的VNTR与IL-1Ra的表达水平有关,因此也与骨质疏松症相关联。
基因结构
IL1RN全称为interleukin 1 receptor antagonist(白细胞介素1受体拮抗物),位于染色体2q14.2,全长16,123 bp。mRNA长度根据其异构体而定,分别为1,802、1,865、1,769、1,973nt。编码的蛋白也有四种长度,分别由159、180、177、143个氨基酸残基组成。
其中,编码分泌型蛋白(sIL-1Ra)和细胞型蛋白(icIL-1Ra)的基因都位于2号染色体2q13-14.1的基因复合体中,由4个共用外显子、3个共用内含子和各自特异的第一个外显子组成。sIL-1Ra基因的特异外显子处于第1个共用外显子之前,上面有一个剪接位点。该外显子被剪切后,连接上3种亚型的icIL-1Ra各自特异的外显子,便构成了icIL-1Ra的序列。进一步翻译表达就产生了带信号肽的sIL-1Ra和不带信号肽的icIL-1Ra。
基因分子生物学功能
IL-1具有广泛的生物学作用,例如:①IL-1作为内源性致热原(endogenous pyrogen)而引起炎症反应;②促进T细胞分泌IL-2和表达IL-2R,直接或间接地通过诱导Th2细胞产生IL-4、IL-5、IL-6来促进B细胞增殖和分化,参与特异性免疫应答;③促进单核/巨噬细胞及血管内皮细胞合成细胞因子、粘附分子,激活白细胞粘附作用和合成炎性蛋白;④促进肝细胞合成急性期蛋白(acute phase protein),从而参与炎症反应;⑤促进伤口愈合;⑥刺激造血;⑦直接或间接发挥杀瘤效应;⑧IL-1Ra可与IL-1竞争性结合靶细胞表面IL-1R,从而发挥抑制作用。
IL-1可看作是能影响多种细胞的细胞因子家族,由IL-1α、IL-1β及IL-1Ra组成。IL-1α、IL-1β与白介素-1受体(IL-1R)结合而发挥广泛的细胞活性激动剂的作用[1]。IL-1Ra(编码基因为IL1RN)是80年代从尿中分离出,可拮抗IL-1作用的细胞因子。其氨基酸序列与IL-1β有26%同源性,与IL-1α有19%同源性,能竞争性地结合IL-1R,但不引起目前可测的激动性效应,是天然存在的IL-1拮抗剂。目前已发现的IL-1Ra有2类4型:分泌型(sIL-1Ra)、细胞型(icIL-1Ra)Ⅰ、细胞型Ⅱ、细胞型Ⅲ[2]。sIL-1Ra主要由单核细胞、巨噬细胞、中性粒细胞等分泌,其N端含有一个25个氨基酸残基的信号肽,主要作用是直接调节免疫反应,包括自身免疫反应。icIL-1Ra存在于角质细胞、肠上皮细胞以及视网膜上皮细胞等,其N端只带有7个氨基酸残基而非信号肽,因此无法转运至胞外。icIL-1Ra在胞内缓冲IL-1的作用,并削弱IL-1诱导产生的基因表达作用[3],该蛋白还可在细胞坏死时释放到胞外发挥类似sIL-1Ra的效能[4]。
IL-1R分为Ⅰ型和Ⅱ型,均属免疫球蛋白超家族。Ⅰ型受体广泛分布于T细胞、成纤维细胞、平滑肌细胞和内皮细胞表面,是IL-1的信号传导受体。Ⅱ型受体主要分布于B细胞、中性粒细胞和单核/巨噬细胞表面。两者信号转导方式不同,具体见图1。
一般,在细胞表面有大量的IL-1R,只需要极小量(<10个)的IL-1与IL-1R结合,即可产生显著生物学效应。IL-1Ra与IL-1R结合而不产生效应是因为它缺乏与IL-1受体辅助蛋白(IL-1R2Acp)结合的区域,因此不具有进一步激发信息传导的功能。再者IL-1Ra半衰期很短(6min),且容易与血循环中可溶性IL-1R结合从而降低其他组织器官中IL-1Ra的水平,加上病变部位IL-1浓度可能很高,因此要完全拮抗IL-1的作用需要10甚至1000倍于IL-1的IL-1Ra数量才能有效。
参与的通路
图1 IL-1R的两种受体蛋白介导的信号转导示意图
图2 IL-1和IL-1Ra的作用途径
基因对疾病发生的影响
白细胞介素-1(IL-1)是一类作用很广泛的细胞因子,参与机体的炎症、免疫防御反应、骨髓造血功能调节及几乎包括所有器官系统的自体平衡机制。其中IL-1α和IL-1β是受体激动剂,与机体免疫功能激活密切相关。研究表明,IL-1常参与各种疾病的发生发展,如败血症、自身免疫性疾病等的病理过程。过多的IL-1会加剧病情的恶化,而恶化的病理过程产生更多IL-1,由此形成恶性循环。另外的研究发现,该家族的另一成员——白介素-1受体拮抗物(IL-1Ra),可缓解此过程,打破恶性循环,因而受到极大的重视。
IL-1在骨代谢中具有双向调节作用,低浓度的IL-1可以促进骨质形成并可作为骨合成代谢的一个指标,然而高浓度的IL-1则可促进骨质丢失而成为与骨密度(BMD)成反比的生化指标。IL-1Ra是特异性与IL-1发生拮抗的细胞因子,必然同IL-1在骨质代谢中发生对抗,从而具有抗骨质疏松症的作用。
功能及人群关联性研究显示,IL1RN基因多态性,特别是研究较多的第2号内含子上VNTR,会影响IL-1Ra的表达水平,因而可能在IL-1介导的骨质流失中扮演重要角色。
还没有人评论,赶快抢个沙发