 腾讯登录
腾讯登录VEGFA
| 导读 | VEGFA的全称为vascular endothelial growth factor A,其中文名为血管内皮生长因子A。
血管生成刺激因素包括有一类称为“血管生成因子(antigenic growth factors)”的细胞因子,起着刺激新血管形成的作用。目前已经发现了一些血管生成刺激因子,如血小板源性生长因子(PDGF)、成纤维细胞生长因子(bFGF)、转换生长因子(TGF)、血管内皮... |
VEGFA的全称为vascular endothelial growth factor A,其中文名为血管内皮生长因子A。
血管生成刺激因素包括有一类称为“血管生成因子(antigenic growth factors)”的细胞因子,起着刺激新血管形成的作用。目前已经发现了一些血管生成刺激因子,如血小板源性生长因子(PDGF)、成纤维细胞生长因子(bFGF)、转换生长因子(TGF)、血管内皮细胞生长因子A(VEGFA)等,其中在肿瘤血管生成中起关键作用的是VEGFA。
VEGFA蛋白由肿瘤细胞、巨噬细胞及纤维母细胞等分泌,广泛分布于人体许多组织,如腺体、肺、肝、肾、心肌等,但表达水平极低,其作用仅维持正常血管密度和基本渗透功能,维持营养物质。当出现肿瘤细胞之后,其表达水平一般会大大上升。有研究表明,VEGFA与肿瘤侵染性相关,与肿瘤易感性亦相关。
基因结构
VEGFA基因位于第6号染色体6p12位置,全长16,272 bp,有7种剪接方法。

VEGFA的7种剪接方法
基因分子生物学功能
血管内皮生长因子A(VEGFA)是最主要的血管内皮生长因子(VEGF),为了与其他多种血管内皮生长因子相关蛋白相区别,以VEGFA命名。VEGF家族目前主要包括VEGFA(即VEGF)、胎盘生长因子、VEGFB、VEGFC、VEGFD和VEGFE,其中VEGFA是诱导肿瘤血管形成作用最强、特异性最高的血管生长因子。
VEGFA是具有高度内皮细胞特异性的有丝分裂原,在血管发生和形成过程中起着主要的调控作用。VEGFA之所以具有高度的内皮细胞特异性是由于该蛋白有3个高亲和性的酪氨酸激酶受体(RTKs),分别为VEGFR-1/Flt-1、VEGFR-2/KDR/Flk-1和VEGFR-3/Flt-4。KDR 是血管形成的主要调控分子,具有明显的化学趋化和促分裂作用,与血管岛、血管形成和造血有关;F1t-l主要在内皮细胞排列形成管腔时发挥作用。这两种受体主要表达在内皮细胞上,虽然极少数造血细胞、单核细胞也有少量表达,但只有内皮细胞对VEGFA有应答反应,因此VEGFA是一个特异作用于血管内皮细胞的生长因子。
VEGFA蛋白的结构是由两条肽链通过二硫键构成的同源二聚体,主要由内皮细胞、单核巨噬细胞、成纤维细胞产生,普遍存在于中枢神经系统、生殖系统以及肿瘤组织中。其主要功能包括:①选择性增强血管内皮细胞有丝分裂,刺激内皮细胞增殖并促进血管形成。②升高血管尤其是微小血管的渗透性,使血浆大分子外渗沉积在血管外基质中,为肿瘤细胞的生长和新生毛细血管网的建立提供营养。
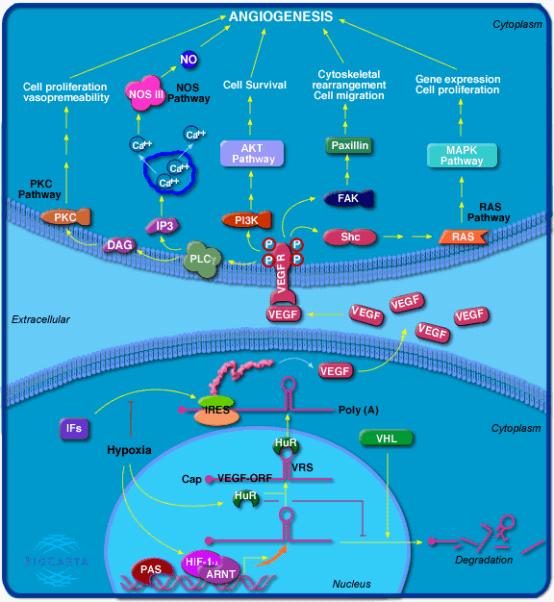
参与的通路VEGFA(VEGF)介导的血管生成通路图
VEGFG在血管生成、肿瘤生长以及缺血性疾病中扮演着重要角色。在体外实验中,缺氧可诱导VEGFA表达。这可能是由于缺氧诱导因子1(hypoxia-inducible factor-1,HIF1)作用于VEGFA基因的5’端的缺氧反应元件,提高了VEGFA的转录效率。生成的VEGFA蛋白与其受体结合,通过PKC、NOS、AKT、MAPK等途径诱导血管生成(图1)。
基因对疾病发生的影响
VEGFA在甲状腺癌的发病过程中起重要作用。它作为血管生长的主要推动因素,可以直接促进甲状腺血管内皮细胞的生长或增加血管的通透性,为成纤维细胞和内皮细胞提供基质,从而促进肿瘤血管的形成。
上世纪70年代,Folkman在《新英格兰医学杂志》中首次提出了肿瘤生长依赖于新血管生成的理论假说。在随后的科学研究中,该假说被进一步证实。肿瘤的生长有两个明显不同的阶段,即从无血管的缓慢生长阶段转变为有血管的快速增殖阶段,血管生成使肿瘤能够获得足够的营养物质,是促成上述转变的关键环节。如果没有血管生成,原发肿瘤的生长不会超过1~2 mm3。肿瘤侵袭转移是肿瘤治疗失败的主要原因,而在肿瘤发生侵袭转移的多步骤过程中,血管生成均发挥着重要作用。
基因功能研究
1989年,Ferrara等人[1]从牛脑垂体滤泡星状细胞中提取到VEGFA,该蛋白是一种肝素结合因子,是由两条N端相同而其他区域存在某些差异的多肽链通过二硫键连接而成的高度保守糖蛋白二聚体,分子量为34-42kD。
1992年,Plate等人[2]通过实验研究后,认为VEGFA可由肿瘤血管内皮细胞表达,在正常内皮细胞组织中无表达。
1994年,Takahashi等人[3]证明VEGFA主要存在于肿瘤细胞,促进肿瘤血管生成,并与其他血管生成相关因子(如胎盘生长因子、碱性成纤维细胞生长因子)之间存在协同作用。
1995年,Freeman等人[4]在对T淋巴细胞的研究中,发现肿瘤浸润淋巴细胞可表达VEGFA。
1997年,Masood等人[5]检测到艾滋Kaposi肉瘤细胞中VEGFA及其受体Flt21和KDR协同高表达,从而首次明确提出肿瘤细胞中VEGFA自分泌作用机制。
1999年,Bellamy等人[6]对造血系统恶性肿瘤(白血病、淋巴瘤、多发性骨髓瘤)的研究发现这些肿瘤细胞不仅表达VEGFA,同时也表达VEGF受体,提示除实体瘤外,在血液系统恶性肿瘤中,VEGFA也可以自分泌途径发挥作用。
2000年,Dias等人[7]证实了VEGFR-2介导的人白血病细胞生长转移的自分泌作用方式。采用人VEGFR-2(KDR)的特异性抗体IMC-C11治疗接种小鼠,可以阻断白血病细胞分泌产生的VEGFA白血病细胞上KDR的相互作用(其与鼠VEGFA体Flt-1无交叉反应),即阻断自分泌作用方式,从而阻碍白血病细胞的生长。并首次证明白血病细胞表达的VEGFA受体是具功能活性的信号转导受体,传递给内皮细胞相似的信号。
2001年,Hayashibara等人[8]亦证实了存在于成人T淋巴细胞白血病中由Flt-1介导的VEGFA自分泌作用机制。并且认为VEGFA及其受体的自分泌作用除了可促进肿瘤细胞生长外,亦可能在肿瘤细胞的转移中发挥重要作用。
2006年,Girnita等人[9]对接受心脏移植的高加索人、西班牙人和美国黑人的16个SNP进行了研究,发现高加索人与西班牙人和美国黑人在多个SNP上存在显著差异。比如,VEGFA -2578CC和-460CC基因型在美国黑人和西班牙人中的分布频率均显著高于高加索人(P<0.001);Fas 的GG基因型和FasL TT基因型常在美国黑人中出现;美国黑人携带ABCB1 2677GG基因型的概率是高加索人的2倍(78.6%比33.7%,P<0.0025)。这样的遗传背景决定了美国黑人更容易发生促炎症和环境因素对其影响更低,以及较低的药效反应和免疫抑制效果。
血管生成刺激因素包括有一类称为“血管生成因子(antigenic growth factors)”的细胞因子,起着刺激新血管形成的作用。目前已经发现了一些血管生成刺激因子,如血小板源性生长因子(PDGF)、成纤维细胞生长因子(bFGF)、转换生长因子(TGF)、血管内皮细胞生长因子A(VEGFA)等,其中在肿瘤血管生成中起关键作用的是VEGFA。
VEGFA蛋白由肿瘤细胞、巨噬细胞及纤维母细胞等分泌,广泛分布于人体许多组织,如腺体、肺、肝、肾、心肌等,但表达水平极低,其作用仅维持正常血管密度和基本渗透功能,维持营养物质。当出现肿瘤细胞之后,其表达水平一般会大大上升。有研究表明,VEGFA与肿瘤侵染性相关,与肿瘤易感性亦相关。
基因结构
VEGFA基因位于第6号染色体6p12位置,全长16,272 bp,有7种剪接方法。
VEGFA的7种剪接方法
基因分子生物学功能
血管内皮生长因子A(VEGFA)是最主要的血管内皮生长因子(VEGF),为了与其他多种血管内皮生长因子相关蛋白相区别,以VEGFA命名。VEGF家族目前主要包括VEGFA(即VEGF)、胎盘生长因子、VEGFB、VEGFC、VEGFD和VEGFE,其中VEGFA是诱导肿瘤血管形成作用最强、特异性最高的血管生长因子。
VEGFA是具有高度内皮细胞特异性的有丝分裂原,在血管发生和形成过程中起着主要的调控作用。VEGFA之所以具有高度的内皮细胞特异性是由于该蛋白有3个高亲和性的酪氨酸激酶受体(RTKs),分别为VEGFR-1/Flt-1、VEGFR-2/KDR/Flk-1和VEGFR-3/Flt-4。KDR 是血管形成的主要调控分子,具有明显的化学趋化和促分裂作用,与血管岛、血管形成和造血有关;F1t-l主要在内皮细胞排列形成管腔时发挥作用。这两种受体主要表达在内皮细胞上,虽然极少数造血细胞、单核细胞也有少量表达,但只有内皮细胞对VEGFA有应答反应,因此VEGFA是一个特异作用于血管内皮细胞的生长因子。
VEGFA蛋白的结构是由两条肽链通过二硫键构成的同源二聚体,主要由内皮细胞、单核巨噬细胞、成纤维细胞产生,普遍存在于中枢神经系统、生殖系统以及肿瘤组织中。其主要功能包括:①选择性增强血管内皮细胞有丝分裂,刺激内皮细胞增殖并促进血管形成。②升高血管尤其是微小血管的渗透性,使血浆大分子外渗沉积在血管外基质中,为肿瘤细胞的生长和新生毛细血管网的建立提供营养。
参与的通路VEGFA(VEGF)介导的血管生成通路图
VEGFG在血管生成、肿瘤生长以及缺血性疾病中扮演着重要角色。在体外实验中,缺氧可诱导VEGFA表达。这可能是由于缺氧诱导因子1(hypoxia-inducible factor-1,HIF1)作用于VEGFA基因的5’端的缺氧反应元件,提高了VEGFA的转录效率。生成的VEGFA蛋白与其受体结合,通过PKC、NOS、AKT、MAPK等途径诱导血管生成(图1)。
基因对疾病发生的影响
VEGFA在甲状腺癌的发病过程中起重要作用。它作为血管生长的主要推动因素,可以直接促进甲状腺血管内皮细胞的生长或增加血管的通透性,为成纤维细胞和内皮细胞提供基质,从而促进肿瘤血管的形成。
上世纪70年代,Folkman在《新英格兰医学杂志》中首次提出了肿瘤生长依赖于新血管生成的理论假说。在随后的科学研究中,该假说被进一步证实。肿瘤的生长有两个明显不同的阶段,即从无血管的缓慢生长阶段转变为有血管的快速增殖阶段,血管生成使肿瘤能够获得足够的营养物质,是促成上述转变的关键环节。如果没有血管生成,原发肿瘤的生长不会超过1~2 mm3。肿瘤侵袭转移是肿瘤治疗失败的主要原因,而在肿瘤发生侵袭转移的多步骤过程中,血管生成均发挥着重要作用。
基因功能研究
1989年,Ferrara等人[1]从牛脑垂体滤泡星状细胞中提取到VEGFA,该蛋白是一种肝素结合因子,是由两条N端相同而其他区域存在某些差异的多肽链通过二硫键连接而成的高度保守糖蛋白二聚体,分子量为34-42kD。
1992年,Plate等人[2]通过实验研究后,认为VEGFA可由肿瘤血管内皮细胞表达,在正常内皮细胞组织中无表达。
1994年,Takahashi等人[3]证明VEGFA主要存在于肿瘤细胞,促进肿瘤血管生成,并与其他血管生成相关因子(如胎盘生长因子、碱性成纤维细胞生长因子)之间存在协同作用。
1995年,Freeman等人[4]在对T淋巴细胞的研究中,发现肿瘤浸润淋巴细胞可表达VEGFA。
1997年,Masood等人[5]检测到艾滋Kaposi肉瘤细胞中VEGFA及其受体Flt21和KDR协同高表达,从而首次明确提出肿瘤细胞中VEGFA自分泌作用机制。
1999年,Bellamy等人[6]对造血系统恶性肿瘤(白血病、淋巴瘤、多发性骨髓瘤)的研究发现这些肿瘤细胞不仅表达VEGFA,同时也表达VEGF受体,提示除实体瘤外,在血液系统恶性肿瘤中,VEGFA也可以自分泌途径发挥作用。
2000年,Dias等人[7]证实了VEGFR-2介导的人白血病细胞生长转移的自分泌作用方式。采用人VEGFR-2(KDR)的特异性抗体IMC-C11治疗接种小鼠,可以阻断白血病细胞分泌产生的VEGFA白血病细胞上KDR的相互作用(其与鼠VEGFA体Flt-1无交叉反应),即阻断自分泌作用方式,从而阻碍白血病细胞的生长。并首次证明白血病细胞表达的VEGFA受体是具功能活性的信号转导受体,传递给内皮细胞相似的信号。
2001年,Hayashibara等人[8]亦证实了存在于成人T淋巴细胞白血病中由Flt-1介导的VEGFA自分泌作用机制。并且认为VEGFA及其受体的自分泌作用除了可促进肿瘤细胞生长外,亦可能在肿瘤细胞的转移中发挥重要作用。
2006年,Girnita等人[9]对接受心脏移植的高加索人、西班牙人和美国黑人的16个SNP进行了研究,发现高加索人与西班牙人和美国黑人在多个SNP上存在显著差异。比如,VEGFA -2578CC和-460CC基因型在美国黑人和西班牙人中的分布频率均显著高于高加索人(P<0.001);Fas 的GG基因型和FasL TT基因型常在美国黑人中出现;美国黑人携带ABCB1 2677GG基因型的概率是高加索人的2倍(78.6%比33.7%,P<0.0025)。这样的遗传背景决定了美国黑人更容易发生促炎症和环境因素对其影响更低,以及较低的药效反应和免疫抑制效果。
顶一下(0) 回复