 腾讯登录
腾讯登录曹雪涛团队再取突破:干预癌症和病毒感染的潜在靶点
| 导读 | IRF3是介导抗病毒先天免疫反应的核心因子,泛素化对IRF3的蛋白酶体周转至关重要。然而,IRF3泛素化对先天免疫的精确激活和及时解决的时空调控尚不清楚。 |
2024年8月8日,海军军医大学/中国医学科学院/北京协和医学院/南开大学曹雪涛团队合作在期刊《Cell Reports 》上发表了题为”The deubiquitinase BAP1 and E3 ligase UBE3C sequentially target IRF3 to activate and resolve the antiviral innate immune response“的研究论文,该研究发现BRCA1相关蛋白-1 (BAP1)和泛素蛋白连接酶E3C (UBE3C)是病毒感染期间控制IRF3稳定性的关键去泛素酶和泛素酶。

https://doi.org/10.1016/j.celrep.2024.114608
背景知识
01
固有信号的有效激活对于宿主对入侵病原体的反应至关重要。干扰素调节因子3 (IRF3)在多种免疫细胞中组成性表达,在抗病毒固有应答中I型干扰素(IFN)的产生中起重要作用。在正常条件下,IRF3以单体形式存在。病毒感染后,IRF3的C端结构域(CTD)被磷酸化,导致其同源二聚化并转位至细胞核,在细胞核内与共激活因子creb结合蛋白CBP/p300形成转录调节复合体,激活I型IFN和IFN刺激基因(ISGs)的转录。IRF3和IFN的充分活化是有效清除入侵病毒所必需的。然而,适当终止IRF3激活和IFN生成对于避免免疫病理(包括ⅰ型干扰素病,如系统性红斑狼疮)也至关重要。因此,IRF3活性的精确时间调控对于有效清除病毒同时避免免疫病理损伤至关重要。
许多翻译后修饰被报道可以调节IRF3的激活,包括磷酸化、泛素化、甲基化、乙酰化、类泛素化和ISG化。泛素化显著影响IRF3的稳定性和活性,影响抗病毒免疫。IRF3的K63多泛素化对于IRF3的激活是至关重要的,因为用K63R突变体取代野生型泛素(Ub)会抑制IRF3的激活。据报道,E3连接酶RAUL (UBE3C [ub -蛋白连接酶E3C])、c-Cbl、TRIM26和RACK可介导IRF3的K48泛素化,导致其蛋白酶体降解,并作为"制动器"停止I型IFN的产生。适当阶段的“制动”有助于炎症和组织损伤的缓解,但过早制动I型IFN会损害宿主的抗病毒免疫。然而,这些E3泛素化酶对IRF3的精确时空调控仍不清楚。目前还没有E3连接酶特异性地参与感染和炎症消退的后期阶段。因此,仍然需要深入了解E3连接酶模块对IRF3稳定性的调节作用。
BAP1介导IRF3在Lys77上的去泛素化
02
为了鉴定k48连接的IRF3泛素化关键位点,研究人员构建了位点特异性的赖氨酸到精氨酸的突变体,其中14个潜在的泛素化位点在人和小鼠之间是保守的。K77R和K87R显著恢复了IRF3的降解,表明这些可能是聚泛素化位点。K77/87R双突变体进一步减弱了IRF3的降解。所有突变体均保留了与BAP1的结合能力。此外,这些IRF3突变进一步激活IFN-β转录和IRF3二聚化。由于Lys77和Lys87位于IRF3的功能性NLS中,研究人员通过共聚焦显微镜评估了K-R突变对IRF3易位的潜在影响。IRF3野生型与K-R突变型的核易位无明显差异。然而,K-A突变完全消除了其核易位,可能是由于精氨酸同赖氨酸一样也是碱性氨基酸,K-A突变破坏了氨基酸的电荷。这也与之前报道的KR77/78NG和RK86/87LQ突变不能诱导IRF3核易位一致。因此,所有这些数据表明,K77和K87是IRF3泛素化和降解的关键位点。
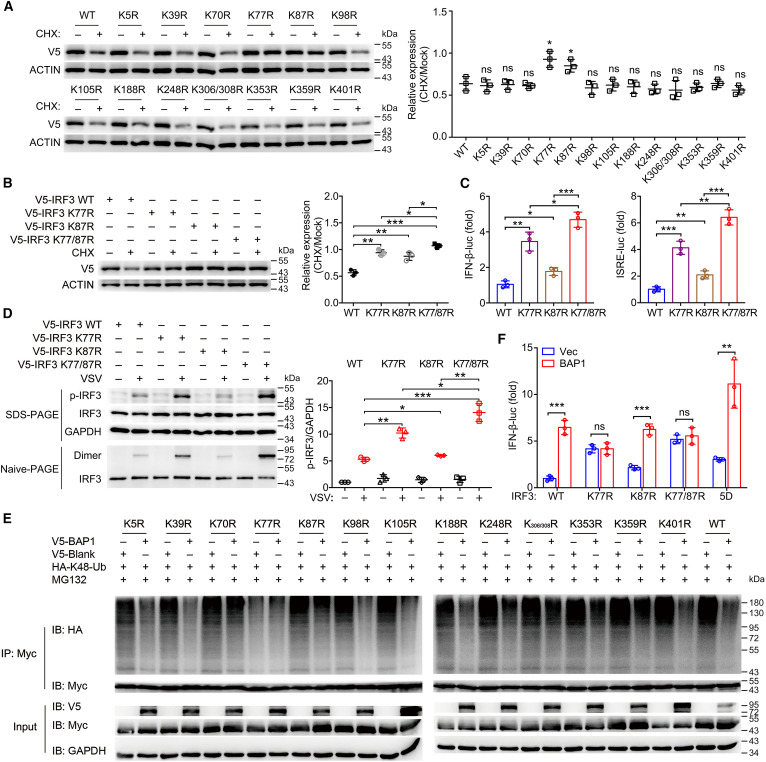
BAP1使IRF3在Lys77上去泛素化
研究人员进一步探索了BAP1靶向的潜在去泛素化位点。BAP1过表达减弱了除K77R外所有突变体中k48连接的多泛素化,这表明BAP1可能特异性介导了IRF3在Lys77上的去泛素化。此外,BAP1增加了野生型IRF3和K87R诱导的IFN-β转录,但在K77R突变体中未观察到这种差异(K77R和K77/ 87r),提示K77位点是BAP1作用的关键。因此,BAP1特异性地介导了IRF3在Lys77上的去泛素化。
BAP1和E3连接酶UBE3C依次结合IRF3并在抗病毒反应中调节Lys77泛素化
03
据报道,几种E3连接酶可介导k48连接的IRF3多泛素化,包括TRIM26、RBCK1、UBE3C和c-Cbl。UBE3C促进k48 -连接的多聚泛素化K87R和野生型IRF3,但不促进K77R和K77/87R IRF3。此外,TRIM26促进k48连接的多聚泛素化的K77R和野生型IRF3,但不促进K87R或K77/87R IRF3。这些结果表明,trim26介导的k48连接的IRF3在Lys87和UBE3C上的多聚泛素化对Lys77泛素化具有特异性。RBCK1和c-Cbl对Lys77和Lys87泛素化均有促进作用,但作用程度较低。
UBE3C在细胞质和细胞核中均有表达,病毒感染后UBE3C在细胞核中富集。而UBE3C在细胞核内特异性地促进了k48连接的IRF3多泛素化。为了进一步明确BAP1和UBE3C在调节IRF3的k48连接多泛素化中的关系,研究人员评估了UBE3C和BAP1在病毒感染过程中的表达动力学。与之相反,UBE3C的表达从感染后12 h开始增加,16 h达到峰值;IRF3的磷酸化在感染后4 h达到峰值,IRF3的二聚体化和核转位在感染后8 h达到峰值,两者均与BAP1的表达一致,但先于UBE3C的表达。这些结果表明,BAP1和UBE3C可能序贯调控k48连接的IRF3多泛素化。为了验证这一假设,研究人员评估了病毒感染期间UBE3C-IRF3和BAP1-IRF3相互作用的动力学。数据显示,BAP1-IRF3(4-12小时)和UBE3C-IRF3(12-20小时)的相互作用在病毒感染过程中被顺序调节。
为了确定BAP1和UBE3C的上调是否依赖于I型IFN的产生,研究人员将野生型、Irf3缺陷型和ifnar1缺陷型小鼠的原代巨噬细胞感染VSV。病毒感染后,Irf3−/−和Ifnar1−/−巨噬细胞中UBE3C的上调消失,而BAP1的上调未见改变,表明I型IFN信号在感染后上调UBE3C。因此,病毒感染后,BAP1和UBE3C可能序贯调控IRF3活性。感染后早期,BAP1表达上调,促进IRF3去泛素化,从而促进抗病毒固有免疫。但在感染后期,I型IFN可能以反馈方式诱导UBE3C表达,并取代BAP1与IRF3相互作用,从而有助于缓解炎症,避免免疫发病机制。
研究结论
04
综上所述,研究人员发现肿瘤抑制基因BAP1可以增强抗病毒固有免疫应答。在机制上,通过去除IRF3 Lys77上的k48连接的多泛素链并防止IRF3被蛋白酶体降解,IRF3信号通路得到加强。本研究结果还揭示了E3泛素连接酶UBE3C和BAP1可以共同调节IRF3的功能。这些发现进一步扩展了研究人员对IRF3表达和功能的调控网络的认识,为肿瘤抑制因子BAP1的生物学功能提供了新的见解,并发现了一个潜在的干预癌症和病毒感染的靶点。(转化医学网360zhyx.com)
【参考资料】
https://doi.org/10.1016/j.celrep.2024.114608
【关于投稿】
转化医学网(360zhyx.com)是转化医学核心门户,旨在推动基础研究、临床诊疗和产业的发展,核心内容涵盖组学、检验、免疫、肿瘤、心血管、糖尿病等。如您有最新的研究内容发表,欢迎联系我们进行免费报道(公众号菜单栏-在线客服联系),我们的理念:内容创造价值,转化铸就未来!
转化医学网(360zhyx.com)发布的文章旨在介绍前沿医学研究进展,不能作为治疗方案使用;如需获得健康指导,请至正规医院就诊。










还没有人评论,赶快抢个沙发