 腾讯登录
腾讯登录瑞金医院俞焙秦团队:HF10抑制胃上皮分化并诱导胃癌
| 导读 | 本研究揭示了GC组织中PHF10的显著上调,与分化水平呈负相关。PHF10成为GC分化诱导治疗的一个有前途的靶点。 |
2024年8月10日,上海交通大学医学院附属瑞金医院俞焙秦团队在期刊《Cancer Gene Therapy 》上发表了题为“PHF10 inhibits gastric epithelium differentiation and induces gastric cancer carcinogenesis”的研究论文。本研究表明,E2F1-PHF10-DUSP5-pERK1/2轴代表了GC细胞分化障碍发展的重要途径,凸显了PHF10作为GC分化诱导治疗的有前途的靶点。

https://www.nature.com/articles/s41417-024-00820-5
研究背景
01
胃癌(gastric cancer, GC)是一种恶性肿瘤,发病率和死亡率都很高。分化障碍的特征,是GC的一个重要特征。研究表明,CDX1的异常表达,是胃上皮分化障碍和癌变发展的重要因素。干细胞相关重编程因子KLF5和SALL4的转录可促进CDX1的上调,通过Wnt通路导致胃上皮肠化生,并促进GC的进展。分化障碍的病因仍然知之甚少,因此,有必要进一步研究分化障碍的复杂分子机制。
团队之前的研究表明,PHF10通过抑制Caspase-3的转录来增强G1/S转化并抑制GC细胞凋亡,表明其在GC中的致癌作用。此外,与高分化GC相比,PHF10在低分化GC中的表达显著升高。PHF10可能在调节GC分化障碍中起关键作用。
已知SWI/SNF(开关/蔗糖不可发酵)染色质重塑复合物在基因表达、调控方面具有重要意义,特别是在癌症干细胞的细胞维持和分化等过程中。 该复合物由Brahma相关基因1(BRG1)相关因子(BAF)和多溴相关BRG1相关因子(PBAF)组成,由具有ATP酶活性的催化亚基、保守的核心调控亚基和可变亚基组成。SWI/SNF复合物由催化亚基BRG1或BRM,以及核心调节亚基SNF5、BAF155和BAF170组成。最近的高通量测序研究,揭示了GC中SWI/SNF复合物成员(如BRG1、BRM、ARID1A和ARID1B)的频繁突变,表明它们参与了GC的发病机制。 此外,PHF10已被确定为PBAF复合物内的转录调节因子,但其在GC肿瘤发生中SWI/SNF复合物形成中的潜在作用,仍不清楚。
细胞分化障碍是致癌的关键步骤。本研究的重点是PHF10在GC细胞分化疾病中的作用,特别是检查其从正常胃粘膜过渡到GC期间的表达模式。采用组织样本验证PHF10表达模式,评价PHF10水平与临床病理参数的相关性,包括GC分化程度和预后。PHF10越来越被视为指导GC分化,诱导治疗的有前途的靶点。
研究进展
02
PHF10在体内抑制GC细胞分化
本研究检查了SGC7901-shPHF10、MKN28-PHF10及其各自对照组植入肿瘤组织中存在的分化标志物。结果表明,PHF10抑制了与胃上皮成熟分化相关的标志物ATP4B、胃蛋白酶原I、GAST、TFF1、TFF2和GKN1的表达,同时上调了与胃癌干细胞相关的CD44、VIL-1和SOX9、胃上皮干细胞和胃上皮祖细胞的表达。这些发现与体外的实验结果一致。
对既往研究的小鼠异种移植物进行HE染色,团队发现SGC7901对照组的肿瘤细胞,呈弥漫性分布和不规则排列。相反,在SGC7901-shPHF10组中,肿瘤细胞整齐排列,形成胃腺管状结构。MKN28-PHF10和MKN28载体产生的异种移植物中的肿瘤细胞,呈弥漫性和不规则分布,缺乏形成腺体样结构的能力。同时,观察到SGC7901-shPHF10细胞在HE染色下表现出胃腺样结构,具有明显的空泡性改变。为了减轻裸鼠中脂肪细胞的潜在混杂因素,证实了胃上皮标志物ATP4B和PGC的存在,而未检测到小鼠脂肪细胞标记物脂联素。这些发现表明,PHF10可能有助于在体内环境中抑制胃癌细胞分化。
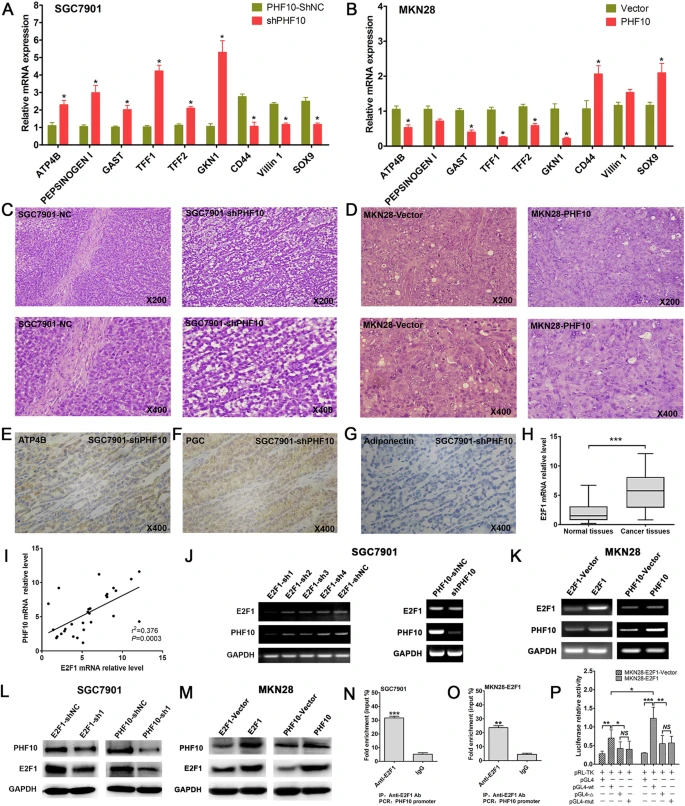
PHF10在体内抑制GC细胞分化,在GC中抑制E2F1-PHF10正反馈。
PHF10通过SWI/SNF复合物直接靶向DUSP5
PHF10是GC细胞分化的重要因子,但其确切的分子机制尚不清楚。鉴于其参与染色质重塑复合物SWI/SNF,据推测,由该复合物介导的转录调控,可能对PHF10在调节GC分化中的作用至关重要。由于TF数据库中缺少PHF10,因此无法直接预测其下游靶点。团队整合和分析了BRG1、SNF5、BAF155和BAF170的ChIP-Seq数据,重点识别与细胞分化相关的得分升高的靶点。研究发现,DUSP5是MAPK(ERK1/2)通路的抑制基因,在启动子的-460~-300 bp区域内有4个结合位点,这些位点与上述分子结合。DUSP5在GC细胞中作为PHF10的下游靶标是合理的,可能会影响细胞分化的调控。
E2F1和PHF10对SGC7901和MKN28细胞DUSP5的表达有负调控作用,而DUSP5对E2F1和PHF10的表达没有影响。敲低PHF10逆转了DUSP5 siRNA引起的DUSP5表达下降,而PHF10的过表达部分,逆转了过表达DUSP5诱导的DUSP5表达增加。随后,ERK1/2的磷酸化水平随着DUSP5的减少或增加,而呈现相应的降低或增加。这与DUSP5作为参与pERK1/2活性磷酸基团去磷酸化的双特异性磷酸酶的既定作用相一致。
BRG1、BAF155和SNF5在使用PHF10免疫沉淀后,使用通用引物,有效扩增了DUSP5的启动子区域。表明SWI/SNF复合物和PHF10的关键组分,能够与DUSP5启动子的同一区域结合。在MKN28-PHF10载体和MKN28-PHF10细胞中转染野生型DUSP5启动子后,荧光显著降低。与野生型组相比,在PHF10结合位点转染截短或突变形式的组,表现出部分逆转的荧光,尽管仍略低于对照组。MKN28-PHF10细胞的荧光强度超过了对照细胞,表明荧光强度与PHF10表达之间存在正相关。这些发现表明,DUSP5是PHF10-SWI/SNF复合物的直接靶标。
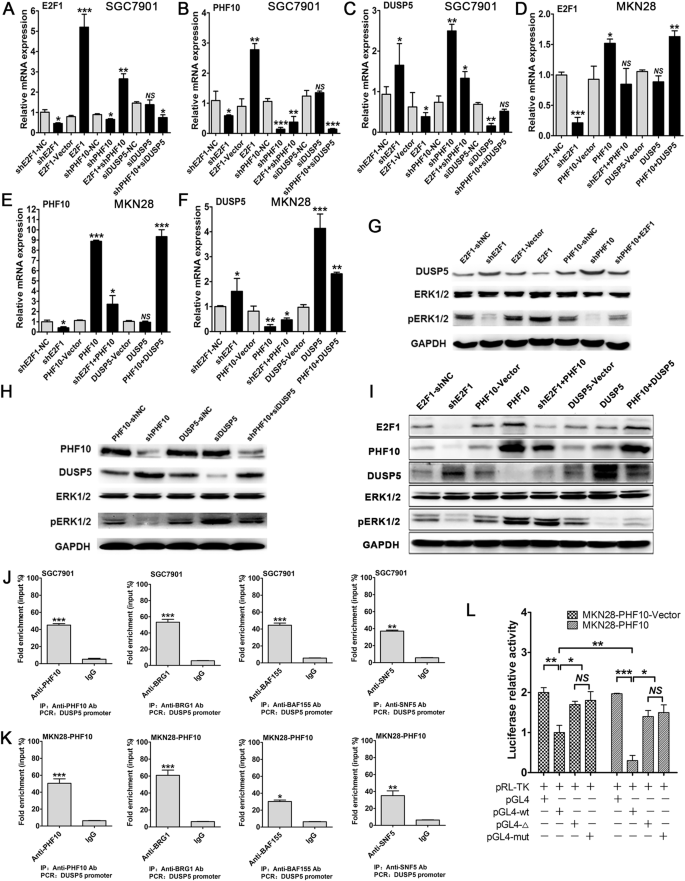
PHF10通过SWI/SNF复合物直接靶向DUSP5。
研究结论
03
尽管GC是分化障碍的常见表现,但其分化失调机制,仍知之甚少。这项研究阐明了涉及PHF10失调的GC细胞分化障碍的关键分子机制。这种失调与GC患者预后密切相关,因为它通过E2F1-PHF10-DUSP5-pERK1/2通路抑制细胞分化,最终促进GC的进展。
PHF10作为TF发挥作用,在介导神经干/祖细胞分化为成熟神经元方面至关重要。 本研究旨在探讨PHF10在GC组织和配对非癌组织中的表达,揭示PHF10表达与分化程度之间存在负相关关系。研究结果表明,从正常胃粘膜到GC进展的不同阶段,PHF10水平逐渐升高。因此,证明了GC的发生、进展和分化异常之间,存在很强的正相关关系。PHF10在体外和体内都抑制了胃上皮细胞的分化,并增强了胃细胞的干性。PHF10在抑制GC细胞分化中的作用,与SWI/SNF染色质重塑复合物密切相关。然而,SWI/SNF 复合物对PHF10调节分化的必要性,需要在未来的研究中通过沉默复合物内的关键组分来验证。
E2F转录因子1(E2F1)是E2F TF家族的一员,在细胞周期调节、凋亡和干性维持等多种生物过程中起着关键作用。本研究旨在探讨气相色谱中PHF10表达失调机制,基于生物信息学预测,E2F1可能作为PHF10的上游TF。团队对GC组织样本的分析显示,E2F1显著上调,其表现出与 PHF10 的共表达。团队通过ChIP-qPCR和双重荧光素酶法,验证了E2F1直接靶向PHF10。PHF10在蛋白质水平上相互调节E2F1的表达,表明两种蛋白质之间存在正反馈回路。这种调控机制可能有助于深入了解GC中E2F1和PHF10同时失调。
双特异性磷酸酶5(DUSP5)是双特异性MAPK磷酸酶(MKPs或DUSPs)家族的一员,以其能够同时水解磷酸化MAPK(ERK1/2、JNK和P38)的丝氨酸和苏氨酸残基上的活性磷酸化基团而闻名,从而将相应的MAPK蛋白恢复到其失活状态。因此,DUSPs作为MAPK通路的负调节因子,起着至关重要的作用。DUSP5表达的抑制与GC患者的不良预后密切相关。本研究进一步阐明,PHF10的过表达通过参与SWI/SNF复合物的组装,促进了DUSP5的直接转录抑制,从而代表了导致GC中DUSP5表达沉默的额外关键机制。DUSP5是PHF10在调节GC细胞分化障碍中的关键靶点。
本研究揭示了信号通路E2F1-PHF10-DUSP5-pERK1/2在GC细胞分化中的重要性。E2F1作为TF发挥作用,直接增强PHF10的转录表达。因此,PHF10还可以提高E2F1的蛋白质水平。这种正反馈机制,导致E2F1和PHF10在GC中的异常过表达。PHF10是SWI/SNF复合物的关键成分,在维持复合物的稳定性方面起着重要作用。此外,团队观察到PHF10与DUSP5启动子相互作用,导致DUSP5的转录抑制。这种抑制导致DUSP5活性降低,特别是在pERK1/2活性磷酸基团的水解中。因此,激活的pERK1/2水平升高,从而激活MAPK通路,并影响细胞功能调节。
参考资料:
1.Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global Cancer Statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71:209–49.
2.Hanahan D. Hallmarks of cancer: new dimensions. Cancer Discov. 2022;12:31–46.










还没有人评论,赶快抢个沙发